

 旧版首页
旧版首页 全新首页
全新首页
 NCS
NCS  学术网站链接
学术网站链接  蛋白质化学工具
蛋白质化学工具  蛋白质平台
蛋白质平台  实用工具网站
实用工具网站 
 学术动态
学术动态  人文建设
人文建设  学术交流
学术交流  人物随感
人物随感  其他新闻
其他新闻  学术周边
学术周边  科学大事件
科学大事件 
 Online PDB
Online PDB  2024年
2024年  2023年
2023年  2022年
2022年  2021年
2021年  2020年
2020年  2019年
2019年  2018年
2018年  2017年
2017年  2016年
2016年  2015年
2015年  2014年
2014年  Prior to 2014
Prior to 2014 
 实验室概况
实验室概况  研究方向
研究方向 
 Thesis Defence
Thesis Defence  Lab Meeting
Lab Meeting  Fun Time
Fun Time  Awards
Awards  Talk
Talk  Notice
Notice  Experiment notebook
Experiment notebook  实验室安全教育
实验室安全教育 
 Groupleader
Groupleader  Associate Professor
Associate Professor  Postdoctoral Fellows
Postdoctoral Fellows  Ph.D Candidates
Ph.D Candidates  Master Candidates
Master Candidates  Undergraduate
Undergraduate  Collaborator
Collaborator  Former Lab Members
Former Lab Members 
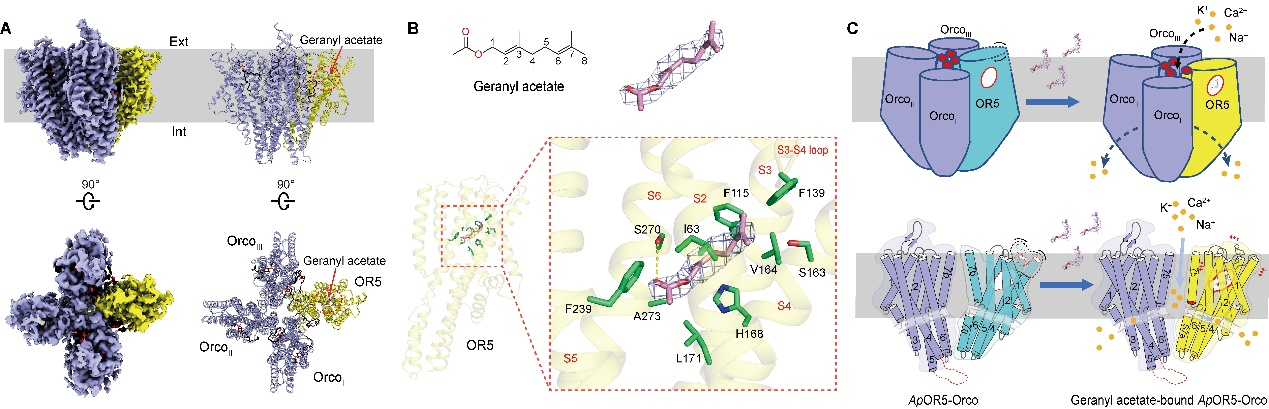

2020年5月12日,Nature Structural & Molecular Biology在线发表了我校作物遗传改良国家重点实验室和生命科学技术学院结构生物学研究团队揭示BIC蛋白抑制蓝光受体隐花色素CRY2激活分子机制的最新研究成果。成果论文以“Structural insights into BIC-mediated inactivation of Arabidopsis cryptochrome 2”为题发表。
光受体是光调控植物生长发育过程中的重要媒介,在整个生长周期中均起到关键作用。隐花色素(Cryptochromes,CRYs)作为蓝光受体参与调控光形态建成如下胚轴伸长的抑制、开花起始、生物节律等重要发育过程。CRYs有两个结构域,包括N端的PHR结构域和C端的CCE结构域。其中PHR结构域非共价结合生色基团黄素腺嘌呤二核苷酸(FAD),对于蓝光的感知至关重要。蓝光激活的CRYs蛋白会产生多种生理生化响应,如FAD光还原、蛋白寡聚和磷酸化等。这些变化有利于CRYs与下游信号因子(转录因子CIBs、 PIFs、BES1等,生长调节因子COP1/SPA复合物等)结合,调控植物生长发育。2016年,加州大学林辰涛教授和福建农林大学王琴教授在拟南芥中鉴定出Blue-light inhibitor of cryptochromes (BICs)蛋白能够负调控CRYs的活性。
在本研究中,作者解析了拟南芥中CRY2蛋白 PHR结构域的晶体结构以及BIC2-CRY2复合物的晶体结构。通过结构比对、分子模拟以及生化实验分析,揭示了BIC2抑制CRY2激活的分子机理。首先,BIC2呈现为极其扩展的结构,蜿蜒曲折的绕着CRY2 以及结构域之间的沟槽,与CRY2形成广泛的氢键和疏水相互作用。这种广泛的互作可能占据了CRY2的寡聚位点从而抑制了CRY2蓝光介导的寡聚化,也能够促进CRY2寡聚体的解聚,使其保持在单体的状态;并且,BIC2可以阻断CRY2寡聚体与下游转录因子CIB1的相互作用从而阻断蓝光信号的传递。其次,BIC2抑制了CRYs蛋白中生色分子FAD的光还原。在光还原过程中,FAD接受外界电子变为FAD•-,CRY2的D393提供一个质子使FAD•-变为FADH•。通过分子模拟发现,BIC2的结合使电子及质子传递的路径变长,从而阻碍电子及质子传递,抑制FAD的光还原。此外,通过序列比对也发现不同植物物种中的BICs与CRYs蛋白互作的区域非常保守,预示这种抑制机制在植物中普遍适用。本研究工作首次解析了植物蓝光受体CRYs与互作蛋白的复合物结构,为研究植物蓝光信号接收传递及调控的分子机制提供了一定工作基础,也为CRYs的光遗传学操作应用提供了一些新思路。
马羚博士、王祥博士和研究生官泽源为本论文共同第一作者,殷平教授为论文通讯作者。校级蛋白质平台为该研究的开展提供了强有力的支持。上海同步辐射光源(SSRF)为数据收集提供了帮助。该研究受到了科技部基金、国家自然科学基金和华中农业大学科技自主创新基金的资助。
文章链接: https://www.nature.com/articles/s41594-020-0410-z
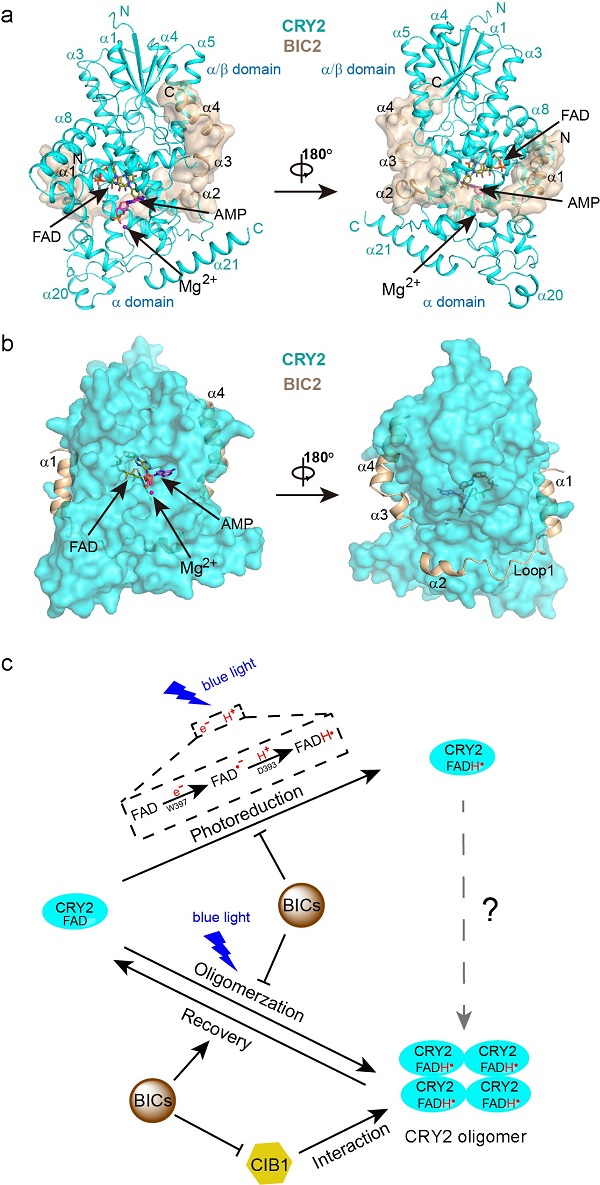
图1 BIC2-CRY2复合物的结构及BICs抑制CRY2的分子机理。
 鄂公网安备 42011102000808号
鄂公网安备 42011102000808号 
